Posted on behalf of Clive Brasier, Emeritus Mycologist with Forest Research, UK
(http://www.forestry.gov.uk/fr/infd-8mraqc)
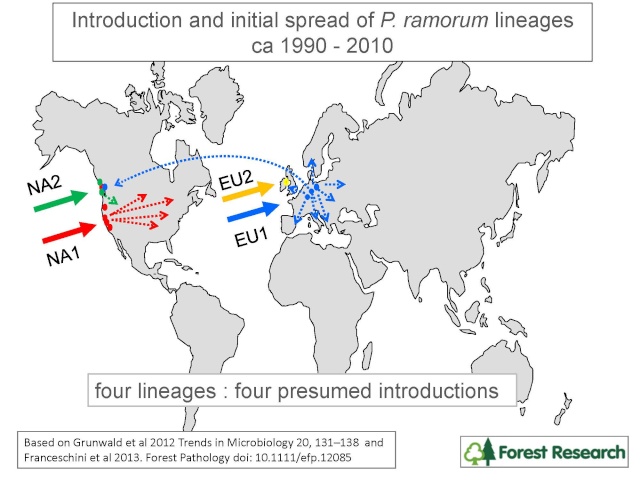
Figure 1. • NA1 was probably introduced into the San Francisco Bay Area in the 1990s. From there it has spread widely into western coastal forests. Its major hosts include tanoak, coast live oak and California bay laurel. NA1 also been distributed across North America within the ornamental nursery trade.
• NA2 was probably introduced into the west coast at about the same time as NA1 but has so far remained primarily confined to a few ornamental nurseries in the Pacific Northwest.
• EU1 was probably introduced into northwest Europe around 1990 and has since been widely distributed in Europe within the nursery trade. In the UK, especially, it has spread from nurseries to wild rhododendron, plantation Japanese larch and other tree species. EU1 has also been introduced into a few North American nurseries, probably from Europe.
• EU2 is the most recently identified lineage. It was probably introduced into Northern Ireland around 2000 and is presently confined to a small area of Northern Ireland and southwest Scotland. It occurs mainly on plantation larch and wild rhododendron.
Speculations on the origins of Phytophthora ramorum•
Phytophthora ramorum was first isolated in Europe in the early 1990s but was not described as a new species of
Phytophthora until 2001. Phylogenetically – from its DNA sequences -
P. ramorum falls into major
Phytophthora Clade 8. Its closest known relatives are
P. lateralis,
P. hibernalis and
P. foliorum. Its geographic origins remain unknown.
• Comparative studies of isolates from North America and Europe have shown that
P. ramorum occurs as four distinct evolutionary lineages, informally labelled NA1, NA2, EU1 and EU2. The prefixes NA and EU stand for North America and Europe respectively.
• Their likely pattern of introduction and spread is summarized in the Figure above.
• Biologically the lineages are rather intriguing. They are phenotypically distinct in their colony patterns, growth temperature responses and pathogenic aggressiveness, suggesting they may be adapted to different environments. Each lineage is of a single sexual mating type (A1 or A2): NA1 and NA2 are of the A2 type and EU1 and EU2 of A1 type. Possibly each lineage lost its ‘opposite’ mating type during its introduction. Inter- lineage (A1 x A2) matings are of very low fecundity and result in genetically aberrant progeny. Comparative genomics suggests the lineages may have diverged from each other 150 – 500 thousand years ago.
• Overall the four lineages therefore appear to comprise ancient, partially reproductively isolated, adaptively different populations. This raises a number of important scientific and regulatory issues.
1. What should the nomenclatural status of the lineages be? Populations that are evolutionarily divergent to a significant degree are often formally taxonomically recognised for scientific communication purposes. Should the lineages, therefore, be accorded the status of separate subspecies of
P. ramorum (or even separate species)? Have they already partially speciated?
2. What should the regulatory status of the lineages be? For example, in Europe the more recently arrived EU2 lineage is apparently more pathogenic (aggressive) than the now widely distributed EU1. Should this be recognised at the plant health regulatory level? For example, should specific measures be taken to restrict the spread of EU2 to the rest of Europe and to other parts of the world?
3. Where have the lineages come from? Understanding how the lineages behave in their centers of origin could provide information of value for
P. ramorum disease control and for assessing the risk of importing other unknown tree pathogens. Current information suggests the lineages might have evolved in four different biogeographic locations. Clues to these locations might be obtained :
(a) from climatic modelling, which can narrow down the options. But in practice many maritime locations worldwide appear to be suitable for
P. ramorum.
(b) from the known
P. ramorum host range in Europe and North America (e.g. Ericaceae and Fagaceae). But the ‘true’ host ranges of the lineages in their centers of origin might be very different from their current known host ranges?
(c) from the geographic distribution of
P. ramorum’s close relatives. Thus
P. ramorum’s closest relative,
P. lateralis, was recently discovered in Taiwan. However searches for
P. ramorum in Taiwan (and searches for it in Yunnan and the Himalayas) have so far been negative.
4. Are there yet more lineages of
P. ramorum out there?
Clive Brasier
……………………………………………………………………………………
Disclaimer: The views expressed in this talk are solely those of the author

